SAMHD1(SAM domain and HD domain-containing protein 1)は、ヒトではSAMHD1遺伝子にコードされるタンパク質である。SAMHD1は細胞内酵素であり、樹状細胞、マクロファージ、単球、休止期のCD4+T細胞中でのHIVの複製の遮断を担う。ホスホヒドロラーゼ(リン酸加水分解)活性を示す酵素であり、デオキシリボヌクレオシド三リン酸(dNTP)を無機三リン酸(iPPP)と2'-デオキシヌクレオシド(リン酸基を持たないデオキシヌクレオシド)へ変換する。SAMHD1はこの反応によってdNTPのプールを枯渇させ、ウイルスのcDNA合成の際の逆転写酵素による利用を防ぐことでウイルスの複製を防ぐ。また、SAMHD1はヌクレアーゼ活性も示す。HIV-1の感染制限にはリボヌクレアーゼ活性が必要であることが記載されたものの、近年のデータではSAMHD1による細胞内でのHIV-1の制限にはリボヌクレアーゼ活性は関与していないことが確認されている。
遺伝子
ヒトのSAMHD1をコードする遺伝子は、ヒト樹状細胞cDNAライブラリ中でマウスのIFN-γ誘導遺伝子Mg11のオルソログとして同定された。SAMHD1遺伝子は20番染色体上に位置する。ゲノム上での長さは59,532 bp(chromosome 20:34,954,059–35,013,590)で16個のエクソンからなり、626アミノ酸からなる72.2 kDaのタンパク質をコードする。SAMHD1は細胞周期が進行しているとしていない細胞の双方で発現しているが、抗ウイルス活性は非周期細胞に限定される。
構造
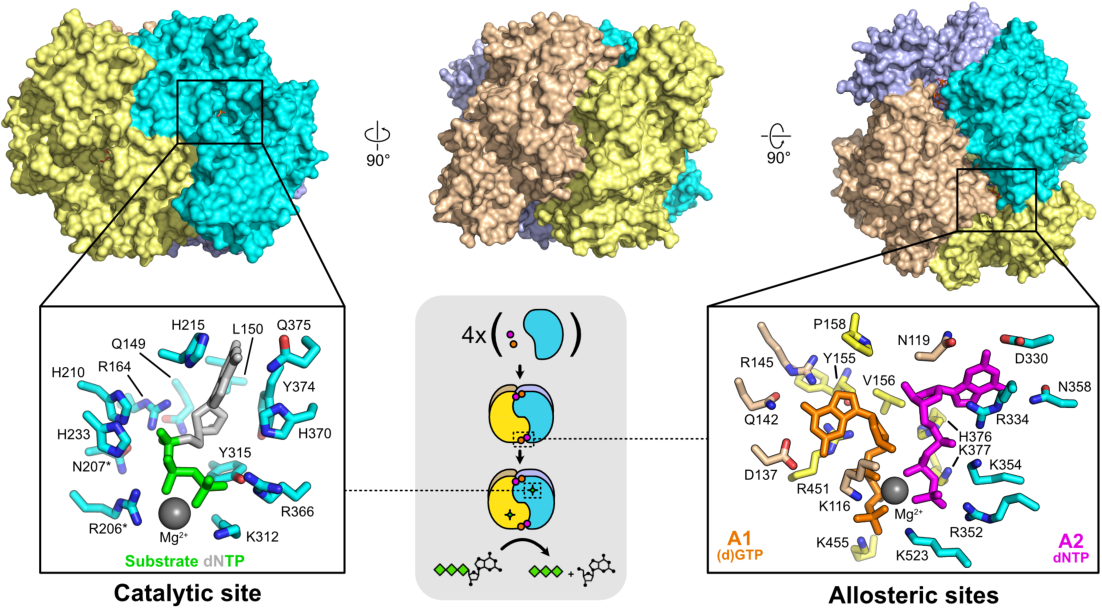
SAMHD1は626アミノ酸からなり、SAMドメインとHDドメインの2つのドメインが含まれる。SAM(Sterile Alpha Motif)ドメインは45–110番残基の領域である。一般的に、SAMドメインは酵母からヒトまで生物においてタンパク質間相互作用やタンパク質-核酸相互作用、キナーゼのドッキング部位、シグナル伝達、転写調節に機能することが知られている。HD(Histidine-Aspartic)ドメインは164–319番残基の領域である。HDドメインを持つタンパク質はヒスチジンとアスパラギン酸からなる触媒残基によって特徴づけられ、dGTPトリホスファターゼ、ホスファターゼ、ホスホジエステラーゼ活性を有することが示されている。
触媒コアを含むSAMHD1断片の結晶構造からは、SAMHD1が二量体であることが明らかにされている。また、SAMHD1はオリゴマー化し四量体を形成することも示されている。SAMHD1は周期細胞ではT592残基がリン酸化されているが、このリン酸化は細胞が非周期状態となった際には失われる。
機能
SAMHD1の変異はエカルディ・グティエール症候群(AGS)でみられる。この疾患はI型インターフェロンの異常な産生によって特徴づけられる遺伝性の自己免疫性脳症であり、その症状は先天性ウイルス感染症と類似している。AGSの患者から単離された単球は、HIV-1に対する感受性が非常に高い。
SAMHD1はレンチウイルスタンパク質Vpxが結合して遮断する宿主タンパク質として同定された。VpxはSAMHD1を標的とすることでマクロファージや樹状細胞への感染を促進する。
ヒトのSAMHD1はdNTPトリホスファターゼ活性、具体的にはdGTP刺激dNTPトリホスホヒドロラーゼ活性と、HDドメインに結合した一本鎖DNAやRNAに対するヌクレアーゼ活性を持つ。他の研究では、SAMHD1のサイレンシングはHIV-1やVpxを欠くサル免疫不全ウイルス(SIV Δvpx)の樹状細胞への感染を高め、また休止期のCD4+T細胞へのHIV-1の感染を高めることが示されている。
疾患における役割
エカルディ・グティエール症候群
エカルディ・グティエール症候群の患者では、SAMHD1遺伝子に16種類の変異が同定されている。変異はSAMHD1の機能を低下させる。しかしながら、このタンパク質の機能不全がどのように免疫系の異常、脳や皮膚への炎症損傷やその他の特徴をもたらしているのかは明らかではない。
ウイルス感染の制限
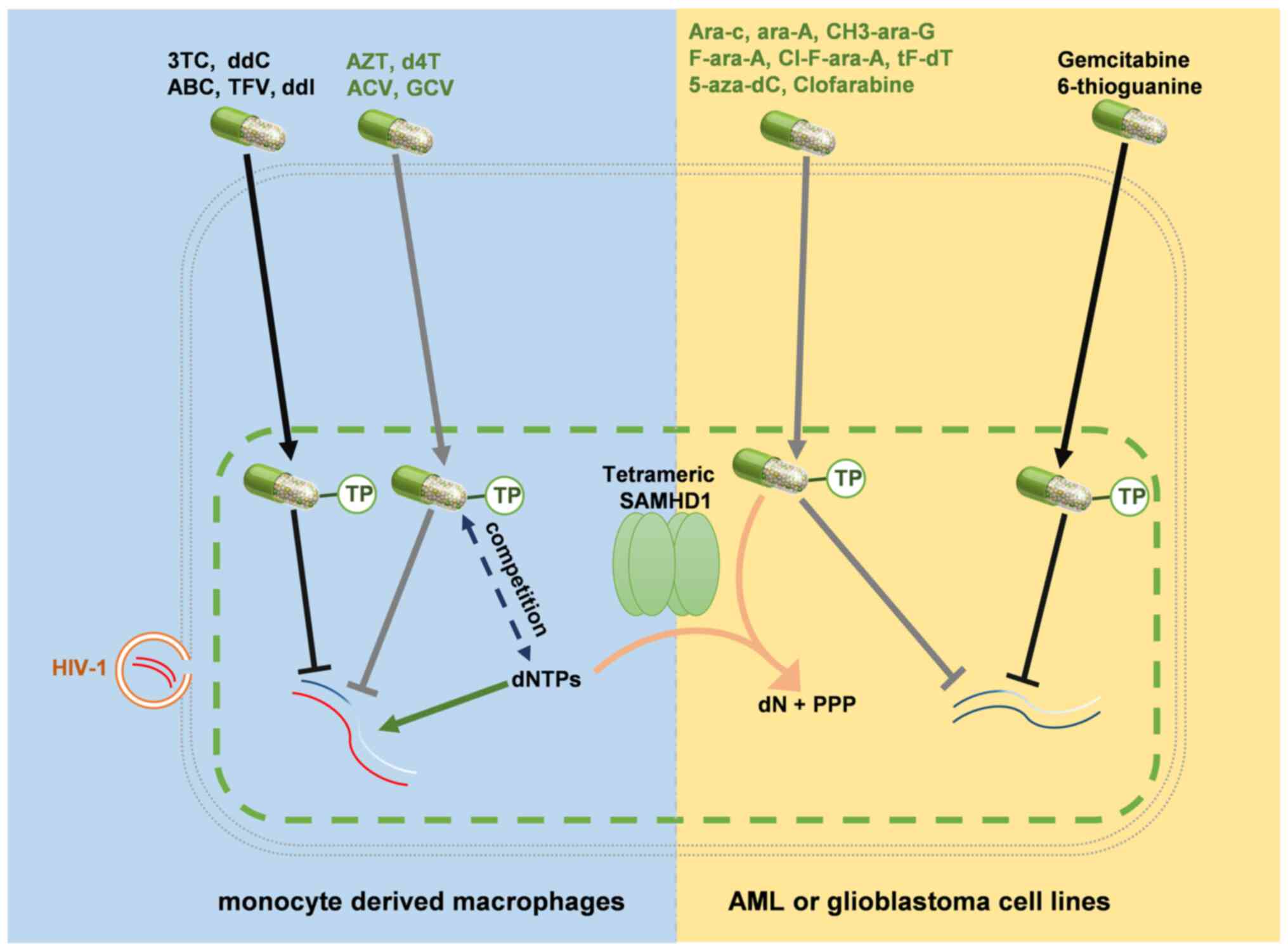
SAMHD1は骨髄系細胞や静止期CD4+T細胞において、HIV-1感染時の逆転写の遮断を担う細胞タンパク質として同定された。骨髄系細胞では、SAMHD1は細胞内のdNTPプールを制限することでHIV-1の感染を阻害する。SAMHD1のdNTPトリホスホヒドロラーゼ活性は細胞内のdNTPレベルを低下させることでHIV-1の感染を防いで免疫系の活性化を防ぐ。SAMHD1のレトロウイルス制限能力はリン酸化によって調節され、このT592のリン酸化はSAMHD1に結合したはサイクリンA2/CDK1複合体によって媒介される。周期細胞でみられるリン酸化されたSAMHD1ははほとんどまたは全く抗ウイルス活性を示さないことが観察されている。逆に、非周期細胞の非リン酸化型SAMHD1は強力な制限活性を示す。
がん細胞における発現調節と代謝拮抗薬分解
がん細胞におけるSAMHD1タンパク質の発現は、4つのレベルで影響を受ける場合がある。まず、SAMHD1遺伝子の変異はSAMHD1のmRNAの産生や、翻訳後の機能的タンパク質の産生を防ぐ場合がある。2つ目に、プロモーターのメチル化もSAMHD1のmRNAの転写を防ぐ場合がある。3つ目に、miRNA-155やmiRNA-181aはSAMHD1のmRNAの翻訳を防ぎ、SAMHD1タンパク質の産生を阻害する場合がある。最後に、細胞周期のS期にはSAMHD1の分解が行われる。B細胞、T細胞、骨髄系細胞は迅速に分裂する場合があり、こうした細胞由来の非接着性の腫瘍細胞株では、接着性細胞と比較してSAMHD1タンパク質は低いか検出できないレベルとなっている。がん細胞におけるSAMHD1によるdNTP濃度の調節は治療との関係において重要な機構である可能性がある。代謝拮抗薬は抗がん剤として利用されるヌクレオシド、ヌクレオチド、塩基アナログであり、いくつかの異なる作用機序で細胞死を促進する。こうした代謝拮抗薬の一部は、アナログの三リン酸型が細胞内で活性化合物として機能する。SAMHD1はアラビノース-5'-三リン酸を加水分解することが示されており、シタラビン(ara-C)応答性に影響を与えるバイオマーカーとなることが示されている。Vpxは血液のがんに対するシタラビン治療の有効性を改善する可能性が提唱されている。
出典
関連文献
外部リンク
- GeneReviews/NCBI/NIH/UW entry on Aicardi-Goutières Syndrome
- OMIM entries on Aicardi-Goutieres syndrome
- Overview of all the structural information available in the PDB for UniProt: Q9Y3Z3 (Human Deoxynucleoside triphosphate triphosphohydrolase SAMHD1) at the PDBe-KB.
- Overview of all the structural information available in the PDB for UniProt: Q60710 (Mouse Deoxynucleoside triphosphate triphosphohydrolase SAMHD1) at the PDBe-KB.